

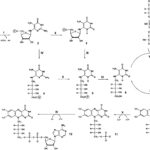
リボフラビンの生合成経路は図1にまとめられているように、1分子のGTPと二分子のリボース5リン酸から始まる(Fischer and Bacher 2005)。GTPシクロヒドロラーゼIIが、アミダゾール環からギ酸、GTP(1)の側鎖から二リン酸を放出して、2,5-ジアミノ-6-リボシルアミノ-4(3H)-ピリミジノーネ5′-リン酸(2)を生成する。2位のアミノ基の水分解性放出とリボース側鎖の還元、そしてリン酸の脱離により5-アミノ-6-リビチルアミノ-2,4(1H,3H)-ピリミジンジオーネ(3)を…

