脂質膜の初期進化と細菌と古細菌の分岐
すべての細胞膜は、グリセロールリン酸のリン脂質でできており、その共通性が最終共通祖先(LUCA)の存在の根拠の一つになっている。しかしながら、リン脂質の生合成経路は、細菌と古細菌で非常に違っているので、それらの最終共通祖先は、リン脂質膜を持っていなかったのではないかとの疑いがもたれていた。Lombard et al 2012によると、最近の系統樹の研究はそれを支持せず、LUCAはむしろ両者の要素を持つ複雑なリン脂質膜を持っていたことを示唆している。
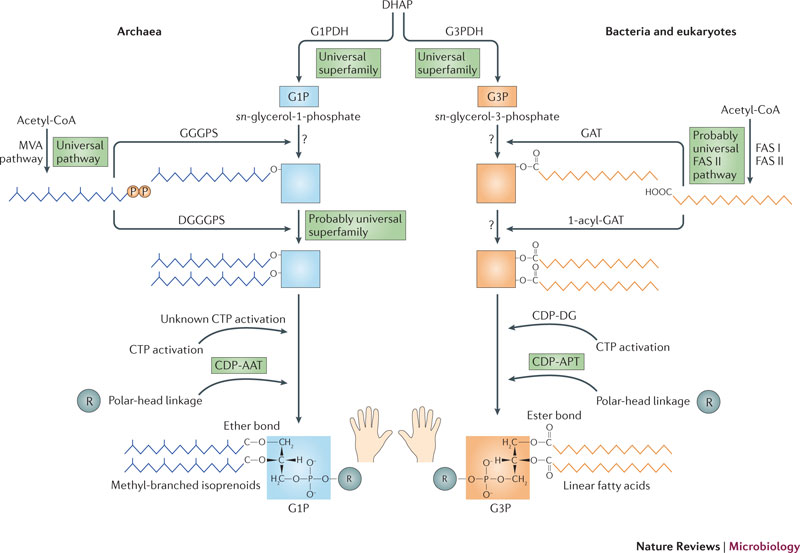
細菌と真核生物は、同じ細胞膜を持っている。それはglycerole-3-phosphate(G3P)でエステル結合された脂肪酸リン脂質でできている。一方で、古細菌の細胞膜はglycerole-1-phosphate(G1P)でエステル結合されたイソプレノイド鎖でできている。細菌と古細菌は鏡映異性的に違う合成経路を使う、特にG1Pを合成するリン酸デヒドロゲナーゼは、G3Pを合成するそれとはまったく違う。このため、LUCAがどのような細胞膜を持っていたかが議論の的になっていた。
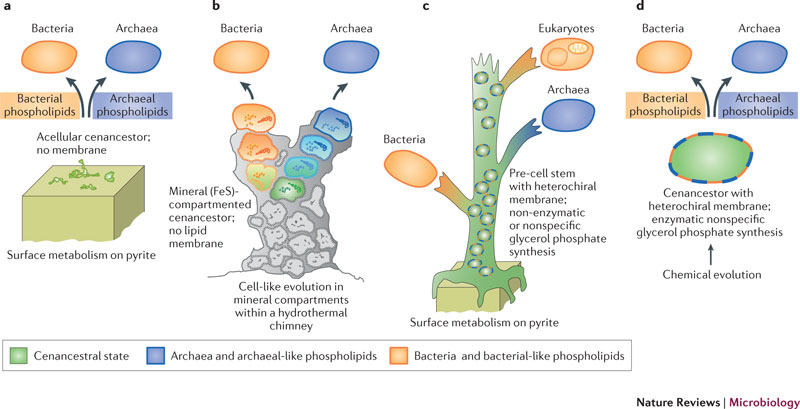
Koga et al. (1998)は、リン脂質生合成経路は、LUCAの後に、細菌と古細菌の系統で別々に確立されたと主張した。しかしこの仮説は、確かに細菌と古細菌の細胞膜の生物の違いを説明するが、LUCAは100種類もの遺伝子を保有しているはずであることが分かってきた。このような複雑なシステムはダーウィン型進化を経て作られると考えられるが、細胞膜で仕切られていない生物がダーウィン進化できる様には思えないので、大変奇妙であった。
Martin and Russell (2003)はリン脂質の代わりに海洋底熱水孔で析出する一硫化鉄鉱物がその仕切りとなり、その内外の酸素濃度、pHと温度の差による化学反応でリボソームとそのほかのLUCAの特質が確立したと考えた。リン脂質の合成系はその後、古細菌と最近の系統で独立に成立したと考える。しかし、海底熱水孔は比較的変化が早く、個別の煙突は典型的には100年以内になくなってしまう。鉱物に束縛された生物は他の煙突への移動が不可能なので、その起源からLUCAまでの進化が一つの煙突内で100年という驚くべき短期間で起こったとは、考えにくい。
教科書は細菌と古細菌のリン脂質の差を強調するが、詳細に調べてみると両者の差はそれほどはっきりしていない。例えば、脂肪酸は古細菌でも検出されており、イソプレノイドも細胞膜の成分としてはすべてのドメインの生命に分布している。唯一の大きな違いは、グリセロールリン酸の鏡像異性体のG1P(古細菌)とG3P(細菌と真核生物)が非相同的なグリセロールリン酸デヒドロゲナーゼを使って行われることである。ただし、最近の遺伝子系統解析は、両者の差の強さについては疑問を呈している。というのは、G1PデヒドロゲナーゼとG3Pデヒドロゲナーゼはどちらも、3つのドメインに広く分布する一つの遺伝子スーパーファミリーに属しているからである。つまりこのスーパーファミリーに属する少なくとも一つの遺伝子は、細菌と古細菌の分岐以前に成立していなければならない。したがって、LUCAはG1PとG3Pを区別せずに合成する祖先的な遺伝子を持っていた可能性がある。その遺伝子が、細菌と古細菌の分岐以後にそれぞれG1PとG3Pに固有な遺伝子に特化していった可能性がある。つまり、LUCAは細菌型のG3P脂肪酸と古細菌型のG1Pイソプレノイドの両方のリン脂質を含んだ細胞膜を持っていたことになる。
最近の系統樹解析は、イソプレノイド合成のためのメバロン酸経路は古細菌と真核生物、そしていくつかの細菌の門で高度に保存していることが分かっている。したがって、メバロン酸回路はLUCAには存在していたが、ほとんどの細菌では失われ、非相同的なメチルエリトリトールリン酸経路にとって代わられたのだろう(Lombert and Moreira 2011)。一方、古細菌は、細菌の脂質合成経路と相同的な遺伝子とを保持し続けた。
では、古細菌と細菌の「細胞膜分岐」はどのように起こったのだろうか。古細菌の最終共通祖先は好熱菌であったことは広く認められている。したがって、古細菌の細胞膜は極端に熱い環境において陽子の細胞膜透過を抑制して細胞膜内外の電気化学的勾配を維持するように進化したのかもしれない(Valentine 2007)。一方、細菌の系統においては、アシル基輸送たんぱく質の進化が、効率的な脂肪酸合成を可能にした。この経路がリン脂質合成系に使われるようになり、イソプレノイド合成系は、他の機能に使われるようになった(Lombard et al. 2007)。脂肪酸もイソプレノイドも、二つの系統の中に残っている。
Lombard et al. 2012, The eraly evolution of lipid membranes and the three domains of life, Nature Review, 10, 507-515.
Koga, Y., Kyuragi, T., Nishihara, M. and Sone, N. 1998, Didarchaeal and bacterial cells arise independently from noncellular precursors? A hypothesis stating that the advent of membrane phospholipid with enantiomeric glycerophosphate backbones caused the separation of the two lines of descent. J. Mol. Evol. 46, 54-63.
Martin, W. & Russell, M. J., 2003, On the origins of cells: a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells. Phil. Trans. R. Soc. Lond. B 358, 59-83.
Lombard, J. & Moreira, 2011, D. Origins and early evolution of the mevalonate pathway of isoprenoid biosynthesis in the three domains of life. Mol. Biol. Evol. 28, 87-99.
Valentine, D. L., 2007, Adaptations to energy stress dictate the ecology and evolution of the Archaea. Nature Rev. Microbiol. 5, 316–323.
Lombard, J., López-García, P. & Moreira, D. An ACPindependent
fatty acid synthesis pathway in archaea? Implications for the origin of phospholipids. Mol. Biol. Evol., 29, 3261–3265

科学はひとつ 宇宙物理学者による知的挑戦の記録
戎崎俊一 著
学而図書/四六判 並製320頁/本体2,400円+税
12年にわたり「戎崎の科学は一つ」で執筆されてきた記事を精選し、「地震と津波防災」など全9章に再編。すべての章に著者書き下ろしの解説を加えて集成した一冊。