内骨格の起源: Origin of endoskeleton
骨芽細胞を含む燐酸カルシウム生物鉱化に関与する細胞は、体表起源だと思われる。
生物鉱化は、まずシアノバクテリアで重炭酸イオン(HCO3-)を光合成に利用するために発達したと考えられる(Arp et al. 2001; Jasson and Northen 2010)。水中では、二酸化炭素溶存量が多くないので、水中光合成生物には、より豊富な重炭酸イオンを炭素源として利用したいという要求が常に存在する。そこで、シアノバクテリアは、以下のような炭素濃縮機構が発達した。まず、重炭酸イオン一個を体内吸収して、体内で
HCO3-+ H+ →H2CO3 → CO2(gas)+H2O
の反応を進め、出てきたCO2(gas)を固定する。この反応が進むためには、体内をアルカリに傾かせない(H+を豊富にする)必要がある。そのために、もしくはHCO3-の代わりに、OH-を体外に放出する。その結果、体外ではアルカリに傾き
Ca2+ + HCO3- +OH- → CaCO3
の反応が進んで炭酸カルシウムが沈殿する。実際、重炭酸イオンを光合成に使う、サンゴやシャジクモでも炭酸カルシウムの沈殿が普遍的に見られる。
シアノバクテリアは、重炭酸イオン利用の老廃物を二次的に再利用し、紫外線や乾燥、外敵から自らを守るための防壁として利用することになったと考えられる。ストロマトライトは、シアノバクテリアを中心とするエコシステムの炭酸カルシウムを主成分とした化石であるとされている。
細胞壁の外側における炭酸カルシウム鉱化機構は、その後大きく進化し、まず、特有の糖たんぱく質が細胞壁の周りに分泌されて集合して、繊維を作り、そこにカルシウムイオンと重炭酸イオンが規則正しく固定される形で、整然と進むようになった。
このような生物鉱化システムの遺伝子は、シアノバクテリア以外の生物にも水平伝播し、多くの生物で利用されるようになった。エディアカラ期からカンブリア期にいたる後生動物の誕生の時期には、炭酸カルシウムの鎧(外骨格)を持った生物が多数誕生した。後生生物は、細菌と共生し、最初はその力を借りて生物鉱化を行っていた。細菌は次第に退化し、その遺伝子は動物に収奪されて平行移動した。このようなイベントは、生物進化上多点で起こり平行な進化を遂げたと考えられる。最近得られた後生動物の系統樹(Peterson and Butterfield 2005)と生物鉱化の分布をみると、生物鉱化は一点で始まったというよりも、多点で始まり平行進化したと考えられる(Murdock and Donoghue 2011)。
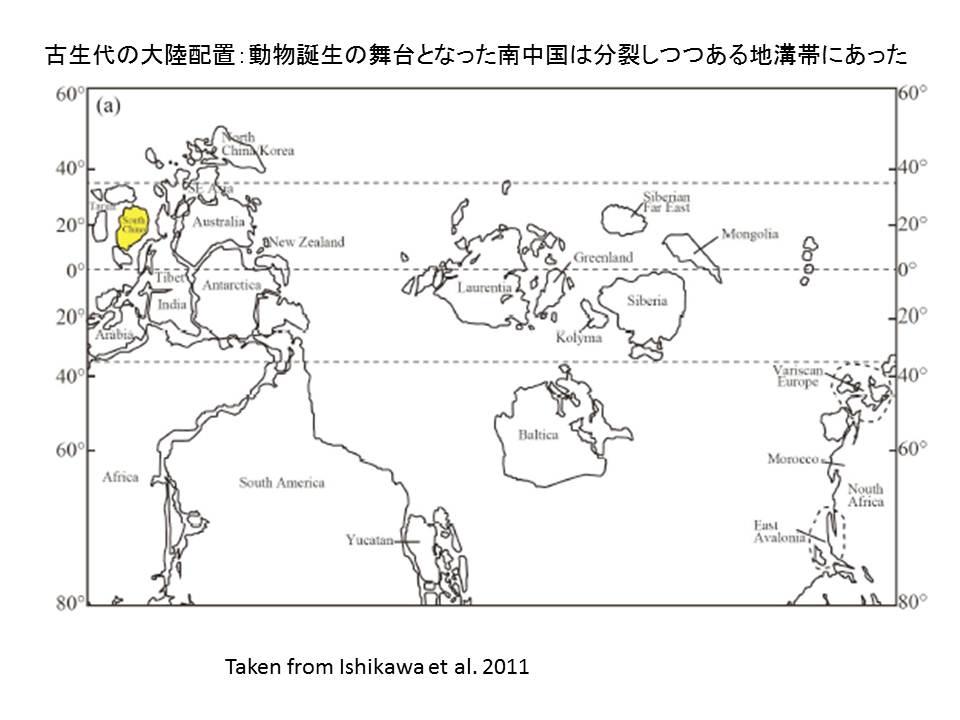
後生動物誕生の舞台となった当時の南中国はオーストラリア大陸と分裂しつつあるリフト地帯に当たっていた。当時は、ロディニア超大陸がたくさんのフラグメントに分裂しつつある時期であった。リフト地帯においては、燐酸に富むカーボナタイトマグマが特異的に噴出することが分かっている。実際、南中国には燐酸鉱床が広がっている。リフト地溝帯に広がりつつあった熱帯の低地帯/湖沼群の一部は、燐酸に富み、燐酸カルシウムに対しても過飽和になった可能性がある。
このような燐酸が豊富な環境において、燐酸カルシウムを外骨格としてもつ生き物も生まれたと考えられる。燐酸カルシウムの生物鉱化メカニズムは糖タンパクの繊維が先行して形成され、カルシウムイオンの固定が整然と行われる点など、炭酸カルシウム鉱化メカニズムとよく似ているので、その遺伝子を改良して発達したと考えられる。
地溝帯がさらに開いて外洋につながると、この燐酸に富む領域は消失した。この結果、燐酸カルシウムを外骨格としてもつ生物は、その海水への溶解を避けるために、それを体内に取り込んで内骨格とするか、表面を粘膜やタンパク質で覆って鱗とするように進化したものと考えられる。
Takemoto et al (2011)は、マウスの体部においては、神経系と中胚葉(骨や筋肉)とが共通の前駆体である“体軸幹細胞”から生まれることを示した。これは、神経系が外胚葉を、骨や筋肉が内胚葉を起源とするこれまでの常識を覆すものである。左右対称動物においては、神経系は外胚葉が陥没してできた神経管から生まれる。骨や筋肉となる細胞が同じ外肺葉起源の幹細胞から生まれる事実は、まず外骨格用として始まった燐酸カルシウムの生物鉱化が、後に内骨格化した進化の事情を反映したものかもしれない。
Jansson C. and Northen T. 2010, Calcifying cyanobacteria-the potential of biomineralization for carbon capture and storage, Current Opinion in Biotechnology, 21, 365-371.
Arp, G. et al. 2001, Photosynthesis-Induced Biofilm Calcification and Calcium Concentration in Phanerozoic Oceans, Science, 292, 1701-1704.
Peterson, K. and Butterfield, N.J., 2005, Origin of the Eumetazoa: Testing ecological predictions of molecular clocks againstthe Proterozoic fossil record, PNAS, 102, 9547-9552.
Murdock and Donoghue 2011, Evolutionary Origins of Animal Skeltal Biominekization, Cell Tissues Organs, 2011, 194, 98-102.
Tatsuya Takemoto, Masanori Uchikawa, Megumi Yoshida, Donald M. Bell, Robin Lovell-Badge, Virginia E. Papaioannou, Hisato Kondoh, Tbx6-dependent Sox2 regulation determines neural or mesodermal fate in axial stem cells, Nature, 470, 394-398 (2011)

科学はひとつ 宇宙物理学者による知的挑戦の記録
戎崎俊一 著
学而図書/四六判 並製320頁/本体2,400円+税
12年にわたり「戎崎の科学は一つ」で執筆されてきた記事を精選し、「地震と津波防災」など全9章に再編。すべての章に著者書き下ろしの解説を加えて集成した一冊。