小型GTP分解酵素の系統樹と真核細胞の進化: Small GTPase and the evolution of the eukaryotic cell
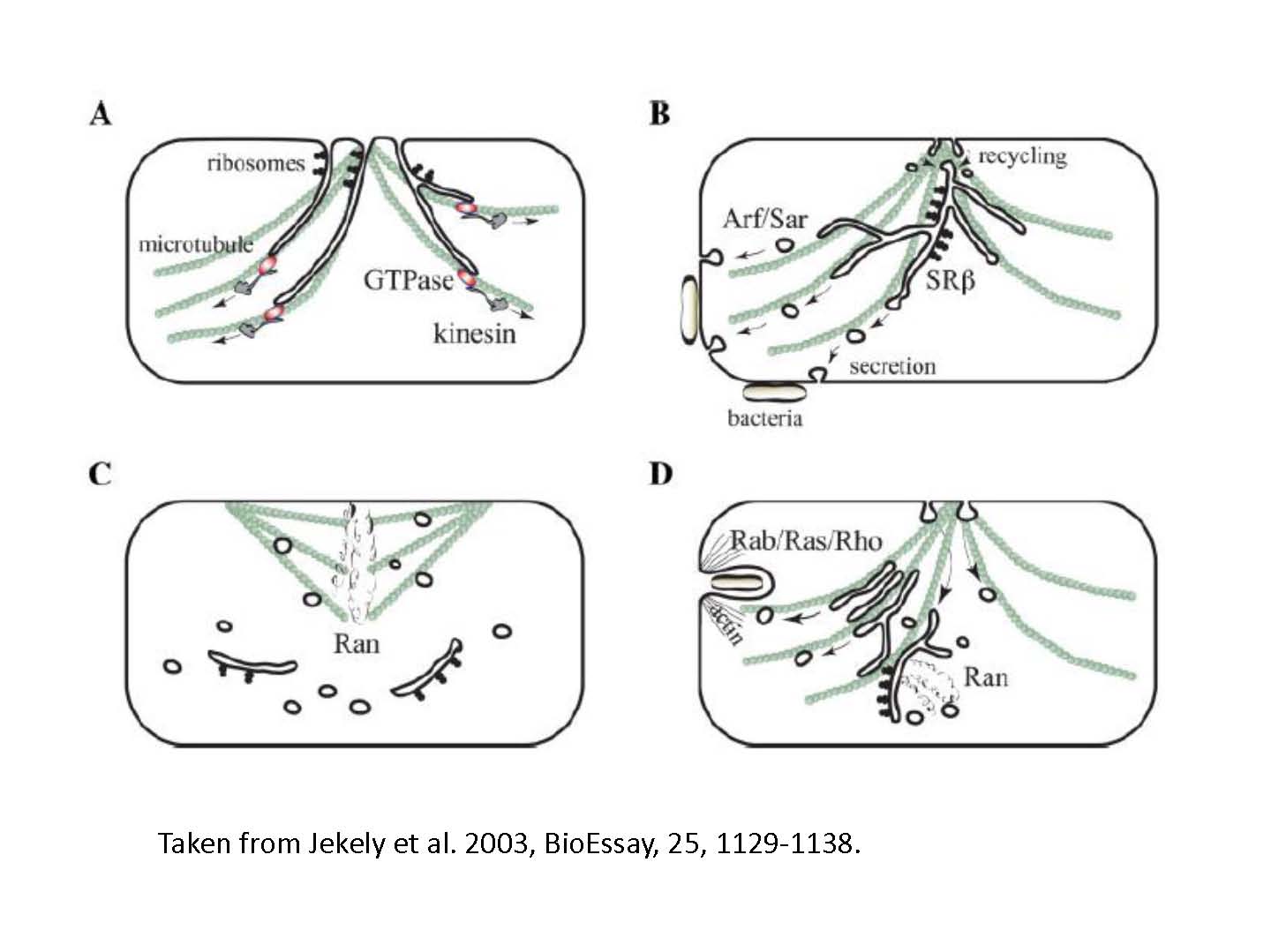
図1:小型GTP分解酵素の系統樹から、真核細胞の進化においてはまず外分泌システムが発達したと示唆された。
RAS超遺伝子族によってコードされる小型GTP分解酵素は、真核細胞において、細胞内骨格ダイナミクス、小胞体運搬、核機能の制御をつかさどる重要な役割を果たしている。Sar1、Arf、SRβ、Rab、Ran、Ras、Rhoを含む小型GTP分解酵素の遺伝子の系統樹を構成してみると、Sar/Arf/SRβ遺伝子族が最初に分岐し、核機能を制御するRan遺伝子族の分岐がその後、食胞機能をつかさどるRab/Ras/Rho遺伝子族はむしろ最後に分岐する結果となった(Jekely 2003)。最初に分岐する。Sar/Arf/SRβ遺伝子族は、分泌性細胞質内膜の分離と融合に関係している。このことを考慮すると、真核生物の進化においては、以下のように、外分泌システムがまず発達していたと推察される。
真核生物の共通祖先(LCEA)は、細菌類が高度に集積したバイオマットにおいて、消化酵素を分泌して近傍の細菌を消化し、分解した栄養分を拡散によって吸収する生活を送っていたと推察されている。そこで、以下のように進化が起こったとJekely(2003)は主張している。
1)放線菌の一種から小型GTPase遺伝子の共通祖先遺伝子を水平移動で獲得した(放線菌は抗生物質をはじめとするいろいろな化学物質を分泌する。LECAは放線菌の一種だったかもしれない)。このGTPaseは細胞膜とキネシンの両方に結合する能力を持っていた。このおかげで微小管に沿って動く分子モーターであるキネシンに引っ張られて、生体膜の管(まだ細胞膜に結合している)が作られるようになった(図1A, Jekely et al 2003)。
2)GTPase遺伝子重複により、Sar、Arf、SRβが分岐し、生体膜の分離と融合が可能になった。これにより細胞膜からは分離した細胞質内膜系が作られるようになった。消化酵素を含んだ小胞体が分離される。それは、微小管に沿ってキネシンで輸送され、細菌の近くで細胞膜に再融合して消化酵素を分泌する。これにより生贄の消化がより効率的になった。食胞機能は持たず、栄養素の吸収はまだ、
拡散に頼っていた(図1B)。
3)さらに遺伝子が重複してRan遺伝子が分岐した。これは細胞分裂の際にDNAと微小管の両方に結合して、その分離を安定化する機能を持った。DNAは、まだ細胞膜に結合しており細胞は細菌と同様の機構で分裂する(図1C)。
4)DNAが結合している生体膜が細胞膜から分離されて、独立の内膜系を構成するようになった。この独立した内膜は、さらにDNAを取り囲んで核膜となりRan遺伝子族がこの制御に当たることになった。さらに、Rab/Ras/Rho遺伝子族が分岐し、アクチン繊維が主要な役割を果たす食胞機能を制御するようになった(図1D)。
真核生物進化の第一人者であるCavaler-Smith2010は、Jekely2003の系統樹の分岐の前後関係は誤差の範囲ではっきりせず、むしろ核の形成、細胞質内膜系、食胞機能の獲得はほぼ同時であったと主張している。
戎崎は、LCEAがバイオマットにすむ肉食生活を送っていたと考えると、まずは外分泌システムを発達させたと考えるのは論理的だと考える。真核生物は、ムレインでできた頑丈な細胞壁を細胞膜の外側に持つ真正細菌ととは違い、N結合糖タンパク質でできた薄くて柔軟な細胞膜しか持っていない。Cavalier-Smithも指摘しているように細胞壁の喪失は、真核生物成立への重要な進化ステップであった。しかし、なぜそれがLCEAにとって有利だったのかが不明だった。もし、LCEAが消化酵素を外分泌して、近傍の細菌を消化し、拡散で栄養を得る生活を送っていたとすれば、細胞壁の喪失によって、消化酵素分泌および栄養素の吸収を早めことができ、有利だったに違いない。
1) Jekely, G. 2003, Small GTPase and evolution of the eukayotic cell, BioEssay, 25, 1129-1138.
2) Cavalier-Smith, T. 2010, Origin of the cell nucleus, mitosis, and sex: role of intracellular coevolution. Biology Direct, 2010, 5, 7.

科学はひとつ 宇宙物理学者による知的挑戦の記録
戎崎俊一 著
学而図書/四六判 並製320頁/本体2,400円+税
12年にわたり「戎崎の科学は一つ」で執筆されてきた記事を精選し、「地震と津波防災」など全9章に再編。すべての章に著者書き下ろしの解説を加えて集成した一冊。