脊椎動物の組織鉱化の進化: Evolution of Vertebrate Tissue Mineralization
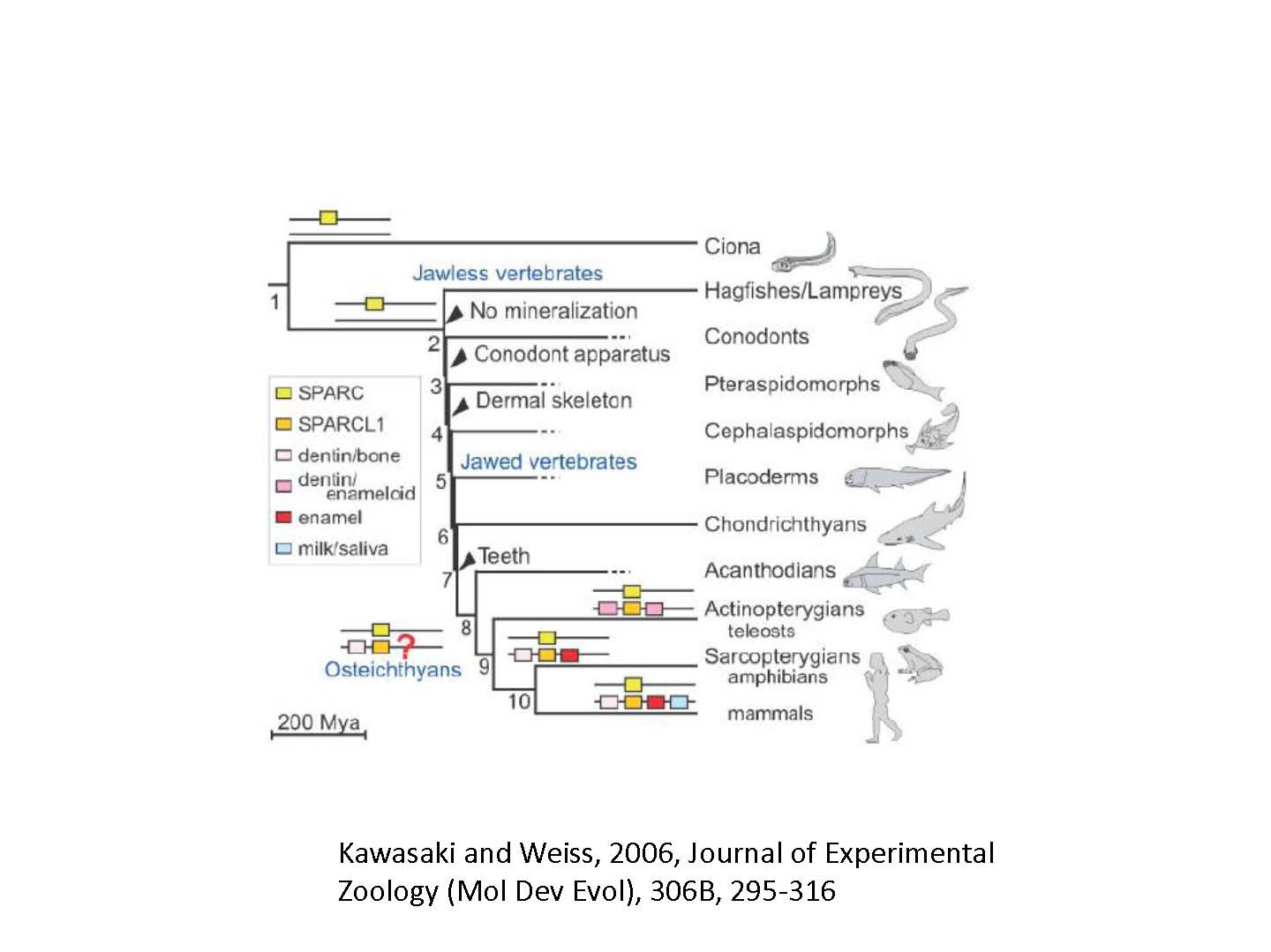
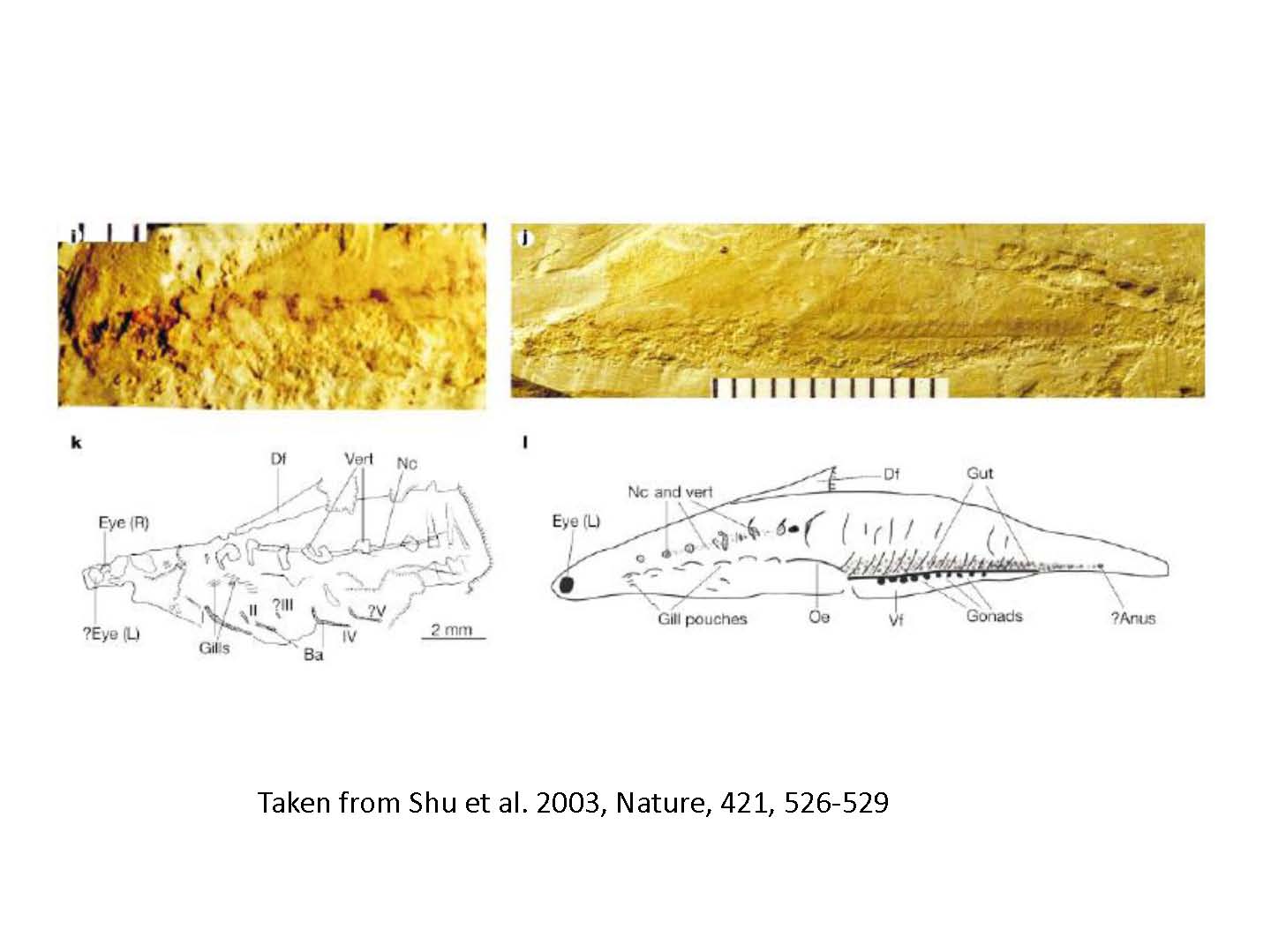
図1左 組織鉱化に多種類の外分泌性カルシウム結合性リンタンパク質(Secretory calcium-binding phosphoproteins: SCPPs)が関与している。哺乳類、鳥類、両生類のこれらのゲノムの相互比較から、SPCC遺伝子の系統樹が作成されその重複の歴史が解明された(図1)。骨/象牙質のSCPPであるSPARCL1、エナメル質のSCPPs(AMBNとANAM)、そしてミルク/唾液SCPPsは、この順でヒトの染色体4に位置している。一方、SPARCは染色体5に位置している。
図2右 カンブリア期の脊椎動物ハイコウイクティスの化石。脊椎と表皮軟骨を持っていたと考えられている。
脊椎動物の組織鉱化の進化: Evolution of Vertebrate Tissue Mineralization
化石の記録を見ると、多くの後生動物がカンブリア期初期(542-513Ma)のカンブリア爆発の時に、鉱化した骨を獲得した。つまり、後口動物、刺皮動物、ホヤは炭酸カルシウムの骨を持つようになった。一方、われわれに近い動物、ナメクジウオと現生の無顎動物、メクラウナギ、ヤツメウナギは鉱化した骨を持っていない。しかし、絶滅した無顎・有顎脊椎動物は燐酸カルシウムの骨を持っている。これらの事実は、脊椎動物の骨は無顎脊椎動物における発明であったことを示している。鉱化した骨を持った最初の脊椎動物はコノドントである。コノドントはカンブリア期初期、もしくは中期に現れ、三畳紀の終わりまで生き延びた(図1ノード3)。彼らは燐酸カルシウムの歯を咽頭口腔部に持ち、食餌器官として機能していた。この歯には象牙質とエナメル質があった。これに次いで、オルドビス期に出現した翼甲類に属する無顎脊椎動物は、装甲を皮膚に発達させていた(図1ノード4)。時にそれは鉱化したエナメル質におおわれていた。現在の有顎脊椎動物の歯は、構造の類似性から、この皮骨に起源していると思われる。
このような組織鉱化には、多種類の外分泌性カルシウム結合性リンタンパク質(Secretory calcium-binding phosphoproteins: SCPPs)が関与している。これらのSCPP遺伝子族は、卵の殻の基質たんぱく質(卵の殻形成)、哺乳類のミルクのカゼイン(授乳の効率化)と唾液のたんぱく質(歯の維持)を制御する重要な遺伝子を含んでいる。哺乳類、鳥類、両生類のSCPP遺伝子の相互比較から、その系統樹が作成され、その重複と分岐の歴史が解明された(Kawasaki and Weiss2006;図1)。
一方、組織鉱化には繊維型コラーゲンも深く関与している。有顎脊椎動物には3つの主要繊維型コラーゲンがある。I型コラーゲンは歯の象牙質、骨、硬骨魚類のエナメロイドの足場に使われ、II型コラーゲンは鉱化軟骨にみられる主要タンパク質である。一方、III型コラーゲンはほとんどのやわらかい組織にみられる。SPARC(secreted protein, acidic, cystene-rich)は線虫から哺乳類までIV型コラーゲンでできたネットワークの基盤薄層に分布している。さらに、SPRACは無顎脊椎動物の表皮に分布してI型コラーゲンの繊維束の成熟化に寄与し、その機械的に強化する。SPARCは酸度が高く、カルシウム結合能力が高いので、脊椎動物の表皮骨形成に使われやすい。
最近の研究によると、二回の全ゲノム重複が、有顎脊椎動物から軟骨魚類が生まれる前に起こったらしい。この二回の全ゲノムバーストで、4つのHox遺伝子クラスタが作られた。同様に、コラーゲンも4つに分岐したらしい。つまり二種類のI型コラーゲン(COL1A1、COL1A2)と、II型(COL2A1)とIII型コラーゲン(COL3A1)がそれである。SCPP遺伝子族は、これらの繊維性コラーゲンの分岐の後に進化したと考えられる。
脊椎動物の最初の組織鉱化は繊維型コラーゲンを足場としSPARCを鉱物結晶化の仲介者として実現したと考えられる。全ゲノム重複による主要繊維型コラーゲンの分岐が最初の組織鉱化をトリガしたかもしれない。SPARCは、前口動物と後口生物の両方に共通にみられるので、SPCC遺伝子族の祖先遺伝子だと考えるのが自然である。その後、条鰭綱、肉鰭綱の分岐の前に、SPARC遺伝子が重複してSPARCL1が作られ、さらに多種類のSCPP遺伝子がこのSPARCL1から遺伝子重複で作られた。特に、硬骨魚類における鉱化組織の進化にはSCPP遺伝子の発展が重要で、その後それらは条鰭綱、肉鰭綱の両方の系統で独立に遺伝子重複が進み、様々な機能を担うようになって現在に至ったと考えられる。
Shu et al. 2003によると、カンブリア期の脊椎動物ハイコウイクティスは、脊索を機械的にサポートする脊椎と頭部に軟骨(半鉱化)を持っていたらしい(図2)。このことと、エナメル質でおおわれた歯を持つコノドントがカンブリア期に出現したことを考え合わせると、上記の脊椎動物の祖先の二回の全ゲノム重複は、カンブリア期以前におこったということになる。カンブリア期の生物の爆発進化には、この脊椎動物の二回の全ゲノム重複が関与したのかもしれない。
1) Kawasaki K. and Weiss, K.M. 2006, Journal of Experimental Zoology (MOL DEV EVOL), 306B, 295-316.
2) Shu, D.-G. et al. 2003, Nature, 421, 526-529.

科学はひとつ 宇宙物理学者による知的挑戦の記録
戎崎俊一 著
学而図書/四六判 並製320頁/本体2,400円+税
12年にわたり「戎崎の科学は一つ」で執筆されてきた記事を精選し、「地震と津波防災」など全9章に再編。すべての章に著者書き下ろしの解説を加えて集成した一冊。