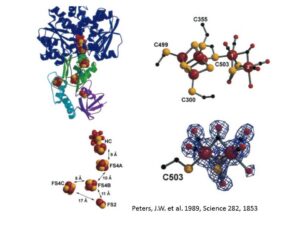
生体におけるATPの形成: Synthesis of ATPs in biological organism
酸化還元反応によって解放されたエネルギーの一部は、高エネルギー結合を持つ生体化学物質に保存される。細胞の中ではアデノシン三リン酸(ATP)というヌクレオチドがエネルギー担体として最も重要である。ATPは、アデノシン塩基とリボース糖一個ずつとリン酸3つでできている。これらのリン酸の酸素原子は、無水結合とエステル結合でつながっている。ATPからアデノシン二リン酸(ADP)へのリン酸の解離でモルあたり50-60 kJのエネルギーが解放される(正確にはリン酸濃度やPHに依存する;Nelson and Cox 2005)。
生きている細胞は、その化学駆動機構を駆動するのに毎秒二百万分子以上のATPを消費する(Purves and Oriants 1983)ので、常に生産されなければならない。これは、光のエネルギーもしくは化学エネルギーを使ってADPに再び燐酸を結合することで達成される。
細胞におけるATP合成は、化学浸透によって行われる(Mitchell 1961)。まず、細胞膜を貫通して分布している様々な電子伝達体が、受け取った電子のエネルギーを使って細胞の中から外に向かって陽子(水素陽イオン)を輸送し、細胞内外に陽子密度の勾配を作る。細胞膜を貫通して存在するATP合成酵素が陽子を細胞内に輸送しつつ、この勾配によってできた陽子駆動力を使って、ATPを形成する。
電子伝達体が膜を通して電子もしくは陽子を輸送して、細胞膜を通した陽子密度の勾配を作る。この過程は、光リン酸化の場合も酸化リン酸化の場合も共通である。普通の細胞においては、以下の6種類の輸送体が働いている(Konhauser 2007)。
1)ヒドロゲナーゼ(hydrogenase)
分子水素 (H2) の可逆的な酸化還元反応を触媒する酵素である。可溶性ヒドロゲナーゼは、細胞質と周辺質に含まれており、水素分子から電子を奪って、NAD(H2+ NAD+-> NADH- + H+)もしくは低い還元状態にある分子(チトクロームc3など)に与える。また、H+からH2の形成を触媒する。
2)フラボタンパク
還元型の補酵素(例えばNADH)や他の分子(例えば乳酸)からH2を取り去る反応を触媒し、H2もしくは電子を次の電子伝達体に渡すことにより、電子伝達鎖を開始する。フラビン・アデニン・ジヌクレオチド(FAD)とフラビン・モノヌクレオチド(FMN)が、フラボタンパクに一種であるNADHデヒドロゲーゼと乳酸エステルのデヒドロゲナーゼの骨格をなしている。
3)鉄・硫黄タンパク
鉄・硫黄クラスタを利用して電子を輸送する。鉄・硫黄反応中心一つからなる単純な分子から多様な鉄・硫黄クラスタを持つ複合体までいろいろある。それぞれの鉄・硫黄クラスタは、少なくとも二つの酸化還元状態、つまり還元型のFe(II)と酸化型のFe(III)を持つ。それぞれのクラスタは、一度に一つの電子を取り扱う。
4)ユビキノン
脂質可溶の輪状構造を持った非タンパク輸送体である。疎水性で電荷をもっていないために、細胞膜の中を自由に拡散する。多様な電子供与酵素から電子を集めて、様々な電子受容酵素に渡す。ユビキノンは、3つの酸化状態を取ることができるつまり、キノン(Q)と呼ばれる最も酸化した状態は、二つの酸素原子を二重結合で環状構造の中に持っている。その一つに水素が付加するとセミキノン(QH・)となり、ハイドロキノン(QH)と呼ばれる最も還元化した状態では、両方の酸素原子に水素が付加されている。
5)チトクローム
鉄を含んだポリフィリン環(ヘムと呼ばれる)を持ったタンパク質である。ポリフィリン環に配位した鉄は、Fe(II)とFe(III)の状態を取ることができる。ポリフィリン環に付加されるサイドグループによって様々な種類のチトクロームが存在する(heme a、b、c、d、そしてo)。それぞれ酸化還元ポテンシャルが違い、したがって、電子伝達鎖の違う場所で使われる。あるものは、電子を次の伝達触媒に渡す還元化触媒として働く(チトクロームc)、またあるものは、最終電子受容体に電子を渡す最終酸化触媒として機能しているものもある(チトクロームaa3)。さらには、電子伝達鎖に外部環境から(S2O32-)電子を供給する役割を持っているものもあり、またその逆(酸化鉄鉱物を還元する)もある。
6)銅タンパク質
Cu(I)とCu(II)の遷移を使って電子を伝達するもので、プラストシアニンとラスチシアニンがある。
呼吸に関与する輸送酵素は、細胞膜の中でその酸化還元電位が最も低いものから高いものへ順に並んでいて、最終電子受容体の還元に使われるまで、段階的にエネルギーが解放されるようになっている。これらは解放されたエネルギーを使って陽子を細胞膜の細胞質側から反対側に輸送する。フラボタンパクやキノンはH2分子を獲得して輸送する。フラボタンパクは、細胞膜全体に貫通していてH2を一方で獲得し反対側で2個の陽子を放出する。ユビキノンは細胞膜の中を自由に端から端まで拡散できる。細胞外へ陽子を放出した後、細胞質内に戻ってそこの水をH+とOH-に分解し、陽子を獲得し、再び細胞外に運ぶ、OH-は細胞質側に残る。一方、鉄・硫黄タンパクやチトクロームは電子のみを輸送する。その場合は、電子が細胞外から中に運ばれ、逆に細胞外にイオンが放出されることにより、差し引きで陽子が細胞外に蓄積する。
電荷を持った陽子(H+)とOH-は細胞膜を通過できないので、細胞膜を通して陽子(そしてOH-)勾配(PHの勾配)と電場が細胞膜を挟んで構成される。ATPaseは、膜を貫通する一種のイオンチャネルで、プロトンもしくは他の陽イオンを通過させる際に解放される陽子駆動力を使ってATPを生産する。
Nelson, D. L. and Cox, M.M. 2005, Lehninger : principle of biochemitry 4th edh. W.H. Freeman, New York.
Purves, w.K. and Oriants, G.H. 1983, Life, The Science of biology, Sinaur, Sunderland, massachusetts.
Mitchell, P. 1961, Coupling of Phosphorylation to electron and hydrogen transfer by a chemi-osmotic type mechanism, Nature, 91, 144-148.
Konhauser, K. 2007, Introduction to geomicrobiology, Blackwell publishing

科学はひとつ 宇宙物理学者による知的挑戦の記録
戎崎俊一 著
学而図書/四六判 並製320頁/本体2,400円+税
12年にわたり「戎崎の科学は一つ」で執筆されてきた記事を精選し、「地震と津波防災」など全9章に再編。すべての章に著者書き下ろしの解説を加えて集成した一冊。