光化学酸化還元反応システムによる前生的な単純糖の形成
前生物的条件における活性化されたピリミジンリボヌクレオチドの合成反応には、酸素的化学反応と窒素的化学反応が混じっている(Powner 2009)。グリコールアルデヒドやグリセルアルデヒドなどの糖の構成要素が、一炭素分子の原料から同様な酸素・窒素混合化学反応系によって作られることが示せれば、RNAが生命の起源にかかわっているという仮説の証拠がより強くなる。Riston and Sutherland (2012)は、これらの糖が、シアン化水素から銅・シアン化物複合体の存在下で、紫外線照射によって作られることを示した。
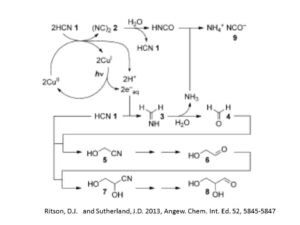
糖はアルカリ水酸化物の存在下でのホルモース反応でホルムアルデヒドが多量体化ことで作られると長年考えられてきた。ホルモース反応では、ホルムアルデヒド1から、グリコールアルデヒド2、そしてグリセルアルデヒド3が合成されるとされている。しかし、カルボニル族の炭素が求電的であって求核的でないので、1→2の二量体化反応には極性変換が必要であるが、それは前生的な環境で存在が示されていないことが問題だ。次の、炭素伸長反応(ホルムアルデヒド1の付加によるグリコールアルデヒド2からグリセルアルデヒド3の生成)にはそれが必要がないので、炭素鎖伸長反応がさらに進みうるが、逆により安定な異性体への変化を止めることができない。このため、余計な反応物ができてしまう。したがって、ホルモース反応は、反応開始体としてグリセロアルデヒドを必要とすることと、反応の制御が困難で複雑な反応生成物ができるのが、RNA合成の観点からは問題だった。
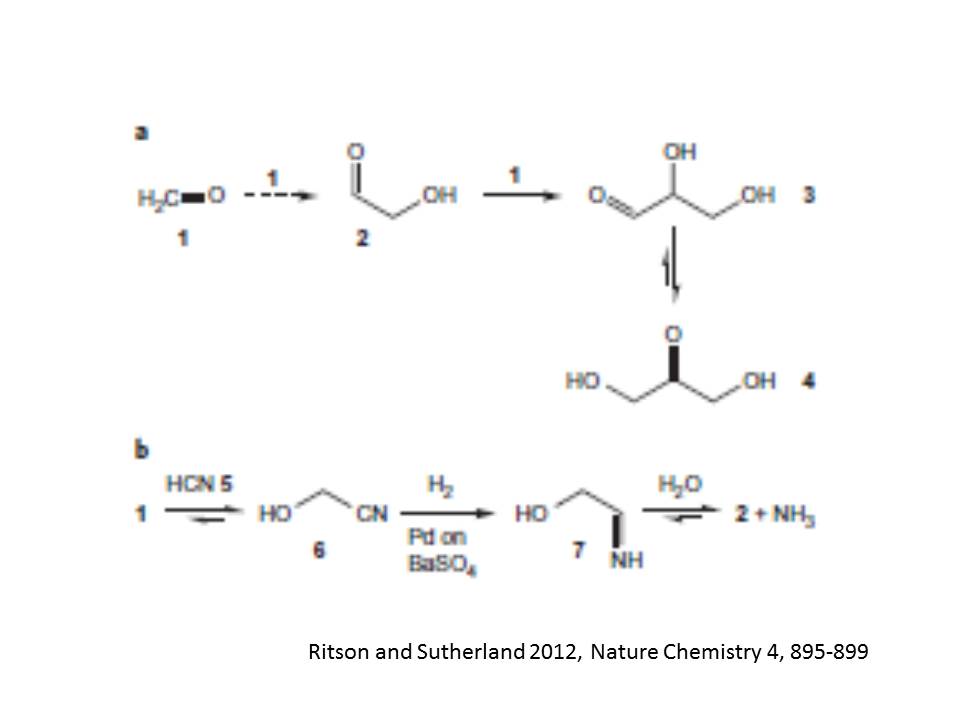
Kiliani-Fischer合成は、ホルムアルデヒド1から始まるもう一つの反復的炭素鎖伸長反応である。この反応系では、シアン陰イオンの求核性を用い、シアンヒドリンもしくはその派生物の形成と、それに引き続く選択的な還元を通して炭素鎖伸長反応を進める。水の中のシアン化水素がシアノヒドリンの一つであるグリコーロニトリル6を作り、それがグリコールアルデヒドイミン7に還元される反応経路が最近見出されている。そのグリセロアルデヒド2とアンモニアへの自発的な加水分解により最初の炭素鎖伸長の第一段階が終了する。ここで、ホルムアルデヒド1とシアン化水素5からシアノヒドリンができる反応は、シアノヒドリンに強く偏った平衡反応(Keq=4.6×10^5 M^-1)であり、運動学的にたいへん容易に進む。その次のグリコールアルデヒドイミン7への反応は、触媒的な水素付加反応による。ただし、グリコールアルデヒドイミン7もしくはグリコアルデヒド2の還元を抑制するための活性抑制触媒(硫酸バナジウムの上のパラジウム)が必須である。でないと、グリコールアルデヒドイミンからグルコルアルデヒドとアンモニアへの平衡分解反応は、酸的な反応条件では、アンモニアのプロトン付加により非加逆になってしまう。普通のKiliani-Fisher反応は、細かく調節された条件を必要とするので、前生的な条件では、有効ではないと考えられてきた。
そこで、Riston and Sutherland (2012)は、光化学反応による水和電子を考慮することにした。一般に、水溶液中の負に帯電した遷移金属シアン複合体は、紫外線照射により、電子を放出する。このようしてつくられた水和電子のニトリルとの相互作用は、これまで化学進化の観点からはあまり論じて来られなかった。
13Cでラベルされたシアン化カリウムと触媒としてのシアン化銅が10%のD2Oが混ざったH2Oの中に溶けている。この水溶液は254nmの紫外線で照射した。試料は随時13C-NMR解析にかけた。7時間後の反応終了時には、NMRスペクトルには、13C-13Cが形成されたことを示す多重信号があった。グリコールアルデヒド2とグリセルアルデヒド3のオキサゾリジン派生物(それぞれ13と14)と思われる。オキサゾリジン派生物は、糖からシアン酸塩との反応で形成されるので、グリコールアルデヒド2とグリセルアルデヒド3が実際に形成されたことを示している。
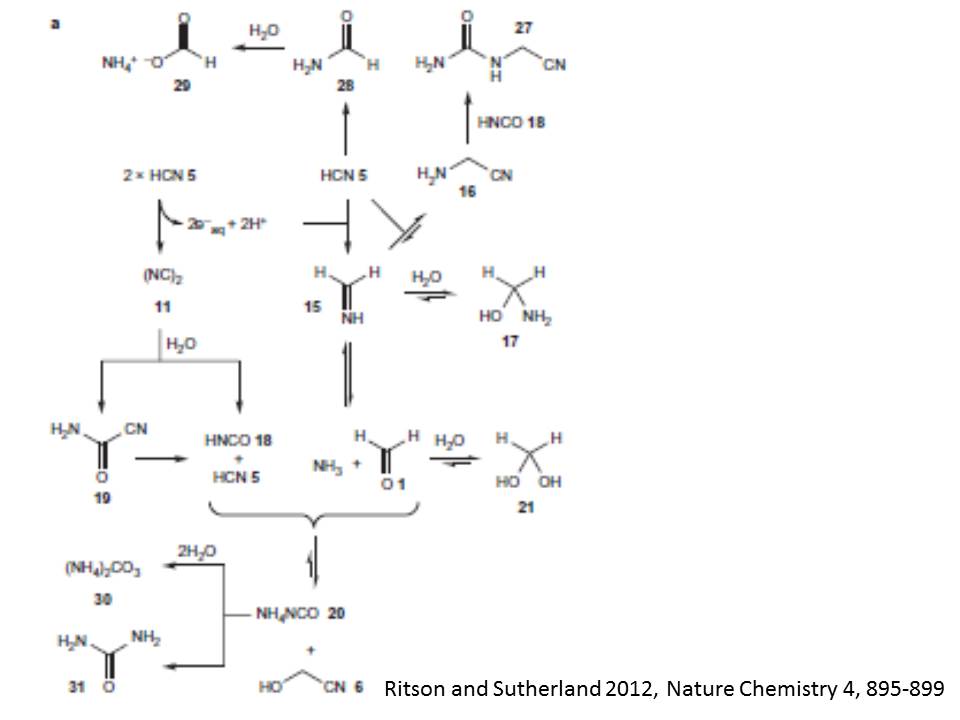
主要な反応過程と中間生成物は以下のように考えられる。まず、反応の第一段階で、シアン化水素5がメタンイミン15に還元される。シアン化水素5が含まれる水中では、メタンイミン15がグリシンニトリル16、ヘミアミナル17、そしてホルムアルデヒド1とアンモニアと平衡状態にある。一方、光酸化還元反応サイクルの生成物であるシアン分子が加水分解して、イソシアン酸18とシアン化水素5の結合への直接・間接の反応経路を与える。生成されたホルムアルデヒド1は、その水酸化物21やグリコールニトリル6との平衡状態にある。
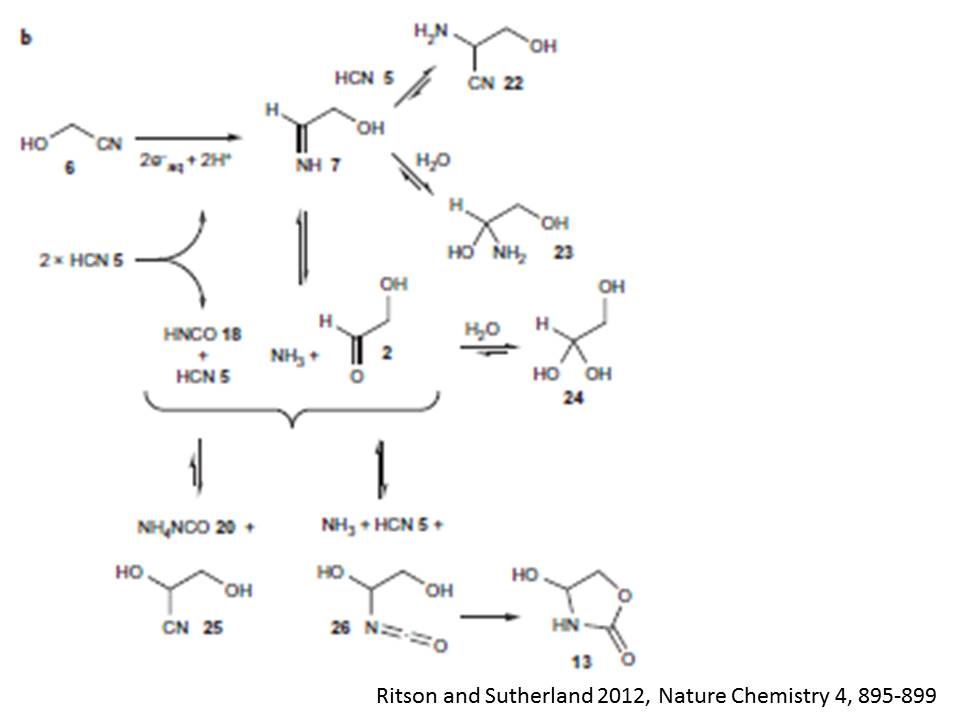
水和電子はシアン化水素5を還元して、上記の第一段階を繰り返すか他のニトリル分子を還元してこれから述べる第二段階を駆動する。グリコールニトリル6の還元は、イミン7をつくる。イミン7はセリンニトリル22とヘミアミナル23、そしてグリコールアルデヒド2とアンモニアと平衡状態にある。アンモニアはイソシアン酸18との酸‐塩基反応で不足するので、この平衡はグリコールアルデヒド2とその水素付加体24、およびシアノヒドリンであるグリセロニトリル25との平衡に置き換えられる。後者の付加反応は平衡状態の非主要成分ではあるが、それはオキサゾリジン環を非可逆的に閉じさせるケミカルポテンシャルを持っている。このオキサゾリジン派生物13は、平衡から離れて蓄積される。さらに、グリセロニトリル25から出発して第二段階反応が進むと、14が形成される。
ヘキサシアノ鉄酸塩のようなシアン金属複合体は光水和や光酸化を進めるが、光酸化還元サイクルについては効率的でない。したがって、シアン化派生物としての隔離なしでの糖の生成が可能であることを証明しなければならない。もし、そのような光化学反応がシアナミドとリン酸の存在下で進行するならば、Powner et al (2008)が示した活性化されたピリミジンヌクレオチドの合成反応と合体が可能になる。
このように、水和電子との反応が生物始原分子同士の反応隘路を広げ、比較的少数の分子同士の反応系においても、相互触媒反応ネットワークの形成を可能とする。ここでは、紫外線の照射による金属シアン複合体の反応によって水和電子を形成しているが、紫外線の代わりに高エネルギー荷電粒子による電離放射線でも同様に水和電子が有効に供給できる。
Riston, D. and Sutherland, J.D., 2012, Prebiotic synthesis of simple sugars by photoredox systems chemisty, Nature chemistry, 4, 895-899.
Powner, M.W. et al. 2009, Synthesis of activated pyrimidine
ribonucleotides in prebiotically plausible conditions, Nature, 459, 239-242.

科学はひとつ 宇宙物理学者による知的挑戦の記録
戎崎俊一 著
学而図書/四六判 並製320頁/本体2,400円+税
12年にわたり「戎崎の科学は一つ」で執筆されてきた記事を精選し、「地震と津波防災」など全9章に再編。すべての章に著者書き下ろしの解説を加えて集成した一冊。